Artículo Original
Caracterización de la enzima similar a trombina del veneno de Bothrops pictus "jergón de costa"
Characterization of thrombin like enzyme FROM Bothrops pictus venom
Dan Vivas-Ruiz1,a, Gustavo A. Sandoval1,a, Fanny Lazo1,b, Edith Rodríguez1,c, Armando Yarlequé1,d, Eladio Flores-Sánchez2,e
1 Laboratorio de Biología Molecular. Facultad de Ciencias Biológicas. Universidad Nacional Mayor de San Marcos. Lima Perú.
2 Laboratorio de Bioquímica y Química de Proteínas, Fundación Ezequiel Días. Bello Horizonte Brasil
a Biólogo, magíster en Biología Molecular; b biólogo, magíster en Biotecnología; c biólogo, magíster en Bioquímica; d biólogo, doctor en Ciencias Biológicas; e biólogo, doctor en Bioquímica
RESUMEN
Objetivos. Realizar una caracterización bioquímica y molecular del principio coagulante del veneno de Bothrops pictus. Materiales y métodos. Se realizó la amplificación del gen a partir de cDNA, se analizó la homología de la secuencia nucleotídica y de la proteína deducida. Se procedió a purificar la enzima para los análisis de secuenciación directa N terminal de los primeros 20 aminoácidos y los ensayos de coagulación sobre plasma humano y fibrinógeno humano, por otro lado, se evaluó el patrón de corte del fibrinógeno por medio de PAGE SDS y la actividad defibrinogenante en roedores albinos (18-22 g). Se determinó el contenido de carbohidratos asociados, el efecto de inhibidores clásicos de proteasas y el efecto de iones bajo la forma de cloruros. Resultados. La enzima mostró homología en la estructura primaria con otras TLEs reportadas para la familia Viperidae, la dosis coagulante mínima (DCM) sobre plasma y fibrinógeno humano fue de 18 y 6 µg respectivamente y su potencia coagulante fue de 131,1 NHI unidades de trombina. La enzima se mostró estable a condiciones fisiológicas y prescinde de iones para su actividad. Los carbohidratos asociados detectados fueron hexosas (25,76%), hexosaminas (13,1%) y ácido siálico (0,76%). Los agentes fluoruro de fenil metil sulfonil floruro (PMSF) ditiotreitol (DTT) fueron los principales inhibidores de la actividad enzimática en tanto que la heparina no tuvo efecto inhibidor. Conclusiones. El principio coagulante del veneno de Bothrops pictus es una enzima similar a trombina.
Palabras clave: Bothrops; Trombina; Coagulación sanguínea; Fibrinógeno (fuente: DeCS BIREME).
ABSTRACT
Objectives. To perform a biochemical and molecular characterization of the coagulant principle from Bothrops pictus venom. Materials and methods. We amplified the genetic sequence of this enzyme from cDNA and analyzed the homology of its nucleotide sequence and its deduced protein. This enzyme was also purified for N-terminal sequencing of first 20 amino acids and for coagulation assays using human plasma and human fibrinogen. Furthermore, cleavage pattern on fibrinogen was evaluated using SDS-PAGE and defibrinogenant activity on white mice (18-22 g). Finally, associated carbohydrate content, effect of protease inhibitors and chloride ions on its enzymatic activity were analyzed. Results. The Thrombin-like Enzyme from Bothrops pictus showed homology at primary level of structure with other previously reported TLEs from Viperidae family. Minimum Coagulant Dosis (MCD) on plasma and human fibrinogen were 18 and 6 µg, respectively, and its coagulant potency was 131.1 NHI Thrombin units. This TLE was stable under physiological conditions and chloride ions are not necessary for its activity. Detected associated carbohydrates were hexoses (25.76%), hexosamines (13.12%) and sialic acid (0.76%). Phenyl methyl sulphonyl fluoride (PMSF) and dithiothreitol (DTT) were the main inhibitors of its enzymatic activity, but heparin had no inhibitor effect. Conclusions. The coagulant principle of Bothrops pictus venom is a Thrombin-like enzyme.
Key words: Bothrops; Thrombin; Blood coagulation; Fibrinogen (source: MeSH NLM).
INTRODUCCIÓN
Los trastornos en el sistema hemostático son el sello característico en el ofidismo, causado por serpientes de la familia Viperidae (jergones); esto se debe a la gama de componentes tóxicos (enzimas y péptidos no enzimáticos) que conforman su veneno y que poseen semejanza en funcionalidad con algunos componentes séricos de la presa o que mimetizan las estructuras de estos conllevando a la activación o inhibición atípica de los procesos de coagulación sanguínea respectivamente (1,2).
Particularmente, existe un grupo de enzimas que actúan sobre el fibrinógeno circulante ya sea degradándolo completamente (enzimas fibrinogenolíticas) o realizando clivajes específicos semejantes a la trombina resultando en la formación de coágulos de fibrina (enzimas coagulantes). Estas últimas reciben el nombre de enzimas similares a trombina (del inglés Thrombin Like Enzyme, TLE) (3). Sin embargo, en su mayoría, las TLE solo liberan un tipo de fibrinopéptido (A o B) y, en muy pocos casos, ambos, lo que produce una malla totalmente inestable y susceptible a degradación por la plasmina (4).
En las serpientes que habitan el Perú, la TLE ha sido identificada en los venenos de Lachesis muta (5), Bothrops bilineatus (6), Bothrops barnetti (7), Bothrops atrox (8) y Bothrops andianus (9). En cuanto a Bothrops pictus, existen reportes de la actividad similar a trombina (10, 11), refiriéndose como una de la más baja del género Bothrops (12-14).
Bothrops pictus, "jergón de costa", es causante de ofidismo en la costa central peruana, su cuadro de envenenamiento se caracteriza por flogosis, eritema, equimosis e impotencia funcional; dentro de los hallazgos de laboratorio la hipoprotrombinemia y prolongación del tiempo de tromboplastina parcial se presenta en la mayoría de los casos (15). Recientemente, nuestro grupo ha reportado la caracterización parcial de la proteína coagulante de B. pictus y en ella se exponen características poco usuales en el grupo de las TLE como mayor rango de pH para su actividad y el incremento de actividad por ion Mn²+ (16).
El interés del estudio de las TLE radica en su potencial aplicación en los campos de diagnóstico clínico y tratamiento de enfermedades cardiovasculares tales como ataques cardiacos, trombosis venosa profunda, infarto agudo de miocardio, trombosis arterial periférica, pérdida repentina de audición neurosensorial así como en la prevención de la formación de trombos tras una intervención quirúrgica (17). Algunas de estas enzimas son ampliamente usadas en los laboratorios de diagnóstico para la detección de fibrinógeno en muestras de sangre heparinizada (17); claro ejemplo de ello son los fármacos Ancrod® y Reptilasa® TLE obtenida de los venenos de Agkistrdon rhodostoma y Bothrops atrox respectivamente (18).
En la presente investigación se evaluó las características bioquímicas de la enzima coagulante del veneno de B. pictus así como su identificación en el ámbito molecular para establecer sus principios funcionales.
MATERIALES Y MÉTODOS
VENENO
El veneno fue obtenido de ejemplares de Bothrops pictus procedentes de la localidad de Pachacamac (región de Lima), mantenidos en el Serpentario Oswaldo Meneses (MHN-UNMSM). Una parte del veneno extraído fue liofilizado para los procedimientos de purificación, mientras que una menor cantidad fue inmediatamente procesado para la obtención del ARN.
OBTENCIÓN Y SECUENCIAMIENTO DEL GEN
La obtención del cDNA y la posterior amplificación del gen fue realizado según Vivas-Ruiz et al.(7), empleando los cebadores F: 5’ ATGGTGCTGATCAGAGTG 3’ y R: 5’ CTGCAATAATGCTCTGGA 3’ diseñados manualmente a partir de alineamiento múltiple de las secuencias batroxobin (J02684.1), bothrombin (AB178321.1), BjussuSP-I (AY251282.1), BITS01A (AF490536.1), Bothrops asper-TLE (DQ2447724.1) y Barnettobin (JX499027). El producto amplificado fue visualizado en gel de agarosa al 1%. El secuenciamiento del gen fue realizado en un secuenciador automatizado ABI 3730 XL (Macrogen, Inc, South Korea). El alineamiento múltiple se realizó empleando el algoritmo del Clustal W del programa BioEdit v 7.2.5; la deducción de la secuencia proteica fue realizada con el programa Translate Tool y la predicción de las propiedades bioquímicas con el programa Protparam ambos programas obtenidos de: http://web.expasy.org/. Tanto la secuencia del cDNA y de la proteína deducida fueron depositados en la base de datos GenBank y UniProt.
PURIFICACIÓN DE LA ENZIMA
Se resuspendieron en buffer acetato de amonio 0,05 M pH 5,0, 100 mg de veneno liofilizado y centrifugados a 2000 rpm. El sobrenadante fue aplicado a una columna de intercambio iónico de CM-Sephadex-C50 (1,2 x 47,5 cm) equilibrado con el buffer antes mencionado y eluído con una gradiente lineal de NaCl de 0,1 a 1 M en el mismo buffer a un flujo 14 mL/h. La actividad enzimática fue monitoreada sobre fibrinógeno y el substrato cromogénico BApNA. Las fracciones con actividad fueron concentradas empleando tubos Microcom. El concentrado resultante (33,9 mg) fue aplicado en una columna de filtración de Sephadex G-100 (1,4 x 64 cm) equilibrada y eluída con el buffer del primer paso, a un flujo de 14 mL/h. Las fracciones activas del paso precedente fueron nuevamente concentradas a un volumen de 0,6 mL (6,4 mg) el cual fue aplicado a una columna de Sephadex G-75 (1 x 30 cm) equilibrada y eluída con el mismo buffer a un flujo de 15 mL/h.
SECUENCIAMIENTO N TERMINAL
La secuencia N-terminal de la enzima fue determinada empleando un secuenciador de proteínas automatizado Shimadzu PPSQ-21A basado en la degradación de Edman, usando una solución con aproximadamente 1 mg/mL de enzima purificada; el procedimiento fue realizado de acuerdo con Magalhaes et al. (19).
DETERMINACIÓN DE CARBOHIDRATOS ASOCIADOS
La presencia de carbohidratos asociados a la proteína fue evaluada mediante la detección de hexosas y hexosaminas de acuerdo a de Winzler R (20) y la presencia de ácido siálico fue evaluada por la metodología de Warren L (21) empleando una solución inicial de proteína a una concentración de 0,5 mg/mL.
ACTIVIDAD ENZIMÁTICA
La actividad coagulante fue medida mezclando fibrinógeno humano (5 mg/mL), en buffer Tris-HCl 0,05M pH 7,4 o plasma humano citratado (0,2 mL) con cantidades apropiadas de enzima (0,5 a 12 µg). Una unidad de actividad coagulante fue considerada como equivalente a una unidad NHI de trombina. La actividad específica fue definida como la cantidad de unidades NHI de trombina por mg de proteína; asimismo, se determinó la dosis coagulante mínima (DCM) (22). La actividad amidolítica fue determinada sobre BApNA a 37 ºC en 50 mM de Tris HCl pH 8,1 siguiendo el incremento de absorbancia a 405 nm a 37 ºC.
ACTIVIDAD FIBRINOGENOLÍTICA
Fue determinada incubando la enzima purificada con 0,1 mL de fibrinógeno 0,2% en buffer Tris HCl 0,05 M pH 7,5 a 37 °C por 10, 20, 30, 60 y 120 min. La incubación fue detenida añadiendo buffer muestra electroforesis y posterior calentamiento a 100 ºC por 3 min. Los resultados fueron visualizados por medio de PAGE-SDS (23).
ACTIVIDAD DEFIBRINOGENANTE
Fue probada en ratones albinos (cepa Balb C 18-22 g) agrupados al azar en seis grupos (cuatro ratones por grupo) a los cuales se les inoculó vía vena caudal dosis decrecientes de la enzima purificada a partir de 50 µg diluidos en 0,1 mL de solución salina. Se determinó la dosis defibrinogenante mínima (DDM) como la mínima dosis de enzima que no produce la coagulación de la sangre total después de 60 min de la inyección intravenosa (22).
EFECTO DEL pH Y TEMPERATURA
Ambos parámetros fueron evaluados sobre BApNA usando 20 µL de la enzima, el efecto del pH se determinó empleando los siguientes buffers:acetato de amonio 50 mM (pH: 4,0 – 6,0); fosfato de sodio 50 mM (pH: 6,0-8,0) y Tris HCl 50 mM (pH: 8,0 -10,0). El efecto de la temperatura se evaluó preincubando la enzima por 15 min en un rango de 4 a 95 ºC.
EFECTO DE IONES E INHIBIDORES
Se probó el efecto de diversos iones bajo la forma de cloruros (c. f.:25 mM) sobre la actividad amidolítica preincubándose con 20 µL de la enzima a 37 ºC por 30 min. Los inhibidores empleados fueron: inhibidor de tripsina de soya (ITS) 1 mg/mL, ácido etilen diamino tetra acético (EDTA), tosil-lisil-clorometil-cetona (TLCK), fenil metil sulfonil flururo (PMSF) y ditiotreitol (DTT) a concentración final de 10 mM. Para el caso de la heparina se probaron tres concentraciones finales: 250, 175 y 87,5 IU (referenciales en la literatura). Estos resultados fueron contrastados usando Trombina bovina (100 IU) como proteína referencial.
ANÁLISIS ESTADÍSTICO
Los datos de actividad enzimática fueron expresados como promedio ± desviación estándar (DS). La comparación estadística de la diferencias fueron realizada empleando ANOVA y el test T de Student. Los valores de p menores que 0,05 (p<0,05) fueron considerados significantes.
RESULTADOS
Se obtuvo una secuencia nucleotídica de 754 pares de bases (GenBank: KF410948) codificante para una proteína de 250 aminoácidos (Figura 1). La proteína madura, denominada como TLE-Bp (Gen Bank: AGZ87932), corresponde a 233 aa con secuencia N-terminal V-I-G-G-D-E propio de las serinoproteasas (4). El peso molecular y punto isoeléctrico determinados fueron 25,9 kDa y 8,2 respectivamente, asimismo, fueron identificados cinco motivos de N-glicosilación.
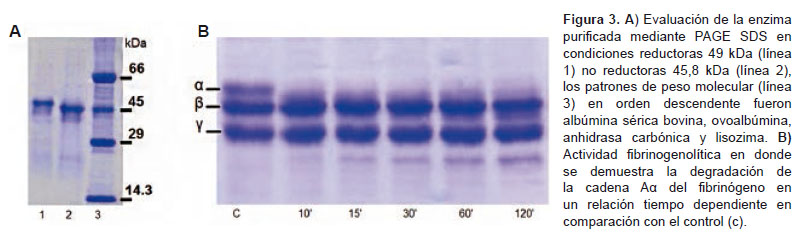
En el análisis por homología (Figura 2) la TLE-Bp muestra regiones conservadas con otras serinoproteasas y enzimas similares a trombina. En estos resultados se destaca la posición de la triada catalítica (His41, Asp85y Ser179) y la disposición de los residuos de cisteínas. La TLE-Bp solo presentó un 30% de homología con la trombina. La TLE-Bp resultó ser una proteína monomérica de 49 kDa (Figura 3a) con una actividad coagulante de 131,1 NHI unidades de trombina sobre Fg humano y actividad amidolítica sobre BApNA. La DCM sobre plasma fue de 18 μg, en tanto que sobre fibrinógeno fue de 6 μg; para ambos caso el coágulo producido por la enzima fue laxo e inestable en comparación de los producidos por la trombina. Los ensayos fibrinogenolíticos evidencia la degradación de la cadena Aα del fibrinógeno (Figura 3b) en tanto que la trombina degradó ambas cadenas, asimismo la DDM fue 1,2 µg/ratón.
Por otro lado, la secuencia de aminoácidos N-terminal de la proteína nativa fue determinada hasta los 20 aminoácidos: VIGGDECNINEHRFLAFTYS (codificación de una sola letra), la cual es congruente con la secuencia proteica deducida. El análisis de carbohidratos asociados demostró que las hexosas son los azúcares mayoritarios (25,76%), seguido por las hexosaminas y el ácido siálico (13,1 y 0,76% respectivamente).
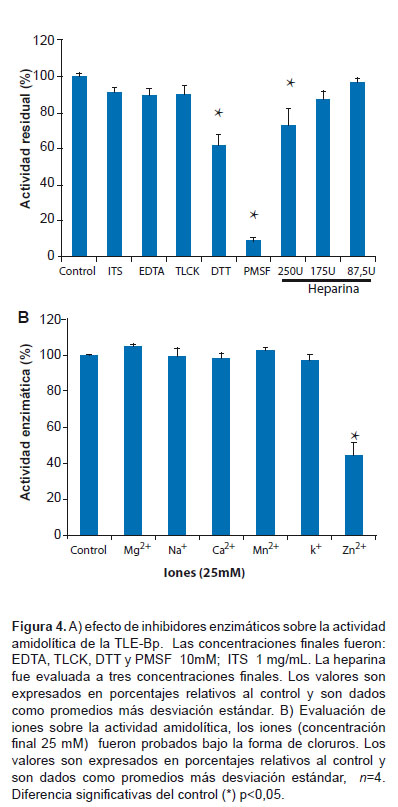
La enzima mostró actividad desde los 30 hasta los 50 oC, la actividad máxima fue reportada a los 35 oC y desde pH 5,0 hasta el pH 11,0, siendo el pH óptimo 8,0. El PMSF redujo la actividad enzimática a 9% e inhibió completamente la actividad fibrinogenolítica, en tanto que el agente reductor DTT y el inhibidor de tripsina de soya la inhibieron a un 65 y 90% respectivamente (Figura 4a). Empleando heparina hasta una concentración de 100 IU/mL no se inhibió la actividad de manera significativa. Por otro lado, los iones Mg2+ y Mn2+ causaron un ligero incremento del 5 y 2% respectivamente, en tanto que el ión Zn²+ causó la reducción en un 52% (Figura 4b).
DISCUSIÓN
Las principales diferencias funcionales de las TLE en comparación a la trombina es el tipo de corte de la molécula de Fg, la mayoría de las TLE cortan solo la cadena Aα o Bβ, no activan el FXIII, coagulan el plasma sanguíneo, pero inhiben la coagulación in vivo, y no son inhibidas por la heparina. Estructuralmente, las TLE son monoméricas con seis puentes disulfuro y solo presentan una similaridad máxima del 30% con respecto a la trombina (4). En la farmacología del envenenamiento por B. pictus se determina alteración de la coagulación (14). Investigaciones previas han reportado la presencia dela actividad coagulante en modelos in vitro e in vivo (13), no obstante, no se ha realizado una caracterización molecular del principio activo responsable.
La estructura primaria de TLE-Bp, determinada por secuenciación directa y deducción del cDNA, muestra el 100% de identidad con la TLE de Lachesis muta y la enzima Ancrod en los primeros 15 aminoácidos, y una gran homología con otras TLE (Figura 2), adicionalmente, con la proteasa A de B. jararaca y una serino proteasa CL4 de Trimeresurus stejnegeri (ambas enzimas fibrinolíticas y no fibrinogenolíticas). La alta homología entre las enzimas coagulantes es confirmada por la presencia del aminoácido valina como el primer residuo N-terminal, una común característica de la mayoría de TLE (24).
La disposición de los aminoácidos pertenecientes al sitio activo (H-D-S) indica que la enzima pertenece a la familia de las serinoproteasas S1 "Clan PA" de endopeptidasas (18), esto se corrobora por la potente inhibición del PMSF el cual se une de manera irreversible al residuo de serina presente en el sitio activo (Figura 4a).
Asimismo, la TLE-Bp mantiene todos los residuos de cisteína en posición conservada con otras TLE (4,20), estos residuos intervienen en la formación de puentes disulfuro de importancia en la estabilidad de la estructura terciaria demostrado por el efecto inhibitorio del agente reductor DTT (4,20). El inhibidor de tripsina de soya no tuvo efecto inhibitorio significativo la cual sustenta una diferencia estructural de la TLE-Bp con esta proteasa.
Por otro lado, la TLE-Bp coaguló el plasma humano y el fibrinógeno humano, sin embargo su inoculación en roedores produjo el efecto de incoagubilidad sanguínea in vivo medida por la actividad desfibrinogenante, este resultado indica que los coágulos inducidos in vivo no poseen la estabilidad optima y podrían ser removidos por los procesos fibrinolíticos endógenos, lo cual produce una depleción significativa del fibrinógeno circulante; la inestabilidad de los coágulos producidos por la TLE-Bp puede estar asociada a la sola hidrolisis de la cadena Aα del fibrinógeno (Figura 3b).
A diferencia de la trombina la TLE-Bp no fue inhibida por la heparina hasta las 100 U/mL, este resultado estaría relacionado a la ausencia en las TLE de ciertas regiones aminoacídicas presentes en la enzima trombina involucrados en la unión de este mucopolisacarido (2,4,5,7). Asimismo, la TLE-Bp no fue potenciada por el Ca2+ ni por otro de los iones evaluados, tampoco es afectada por el agente quelante EDTA; esto indica que la enzima no requiere de iones divalentes para su actividad. Por otro lado, el ion Zn2+ podría ser un inhibidor endógeno de serinoproteasas que evita la autoproteolisis en la glándula productora del veneno (2).
Queda aún por determinar el rol que cumplirían los carbohidratos asociados a la TLE-Bp, debido a que representan aproximadamente el 40% del peso total de la proteína. Adicionalmente, queda por determinar la participación de esta enzima en la alteración de otros procesos fisiológicos durante el desarrollo del envenenamiento.
Nuestros resultados permiten concluir que la enzima coagulante del veneno de Bothrops pictus tiene una actividad similar a trombina y puede ser denominada como tal (denominación Gen bank: Pictobina) ya que posee características semejantes con otras TLE presentes en otras especies.
Agradecimientos: la presente investigación fue realizada gracias al apoyo económico del Programa Nacional de Innovación para la Competitividad y Productividad (Contrato 131—FINCyT-IB-2013) y el convenio CONCYTEC (Perú)-CNPq (Brasil) y forma parte de la Tesis Doctoral de Dan Vivas-Ruiz desarrollada en la Facultad de Ciencias Biológicas-UNMSM.
Contribuciones de autoría: DVR, EFS y AY participaron en la concepción y diseño del artículo. DVR, GAS y ER en la recolección y obtención de resultados DVR y FL participaron en la redacción del artículo. DVR y AY participaron en el análisis e interpretación de datos y aprobación de la versión final. AY, EFS y GAS en la obtención del financiamiento.
Fuente de financiamiento: la realización del presente trabajo ha sido posible gracias a los fondos provenientes del Convenio de Cooperación Bilateral CONCYTEC (Perú) - CNPq (Brasil), así como del Programa Nacional de Innovación para la Competitividad y Productividad - Innóvate Perú (Contrato 131-FINCyT-IB-2013).
Conflictos de Interés: los autores declaran no tener conflicto de interés alguno en la realización y publicación del presente trabajo.
La presente investigación es parte del trabajo de tesis doctoral de Dan Vivas Ruiz.
Referencias Bibliográficas
1. Fry BG, Roelants K, Champagne DE, Scheib H, Tyndall JD, King GF, et al. The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu Rev Genomics Hum Genet. 2009;10:483-511. doi: 10.1146/annurev.genom.9.081307.164356.
2. Stocker K, Fischer H, Meier, J. Thrombin-like snake venom proteinases. Toxicon. 1982;20(1):265-73.
3. Braud S, Bon C, Wisner A. Snake venom proteins acting on hemostasis. Biochimie. 2000 Sep-Oct;82(9-10):851-9.
4. Castro HC, Zingali RB, Albuquerque MG, Pujol-Luz M, Rodrigues CR. Snake venom thrombin-like enzymes: from reptilase to now . Cell Mol Life Sci. 2004 Apr;61(7-8):843-56.
5. Yarlequé A, Campos S, Escobar E, Lazo F, Sanchez N, Hyslop S, et al. Isolation and characterization of a fibrinogen-clotting enzyme from venom of the snake Lachesis muta muta (Peruvian bushmaster). Toxicon. 1989; 27(11):1189-97.
6. Cahuana G, Vivas D, Rodriguez E, Yarleque A. Purificación y características de bilineatobina, una proteína coagulante del veneno de la serpiente peruana arborícola Bothrops bilineatus (loro machaco). Rev Soc Quím Perú. 2012;78(1):43-52.
7. Vivas-Ruiz DE, Sandoval GA, Mendoza J, Inga RR, Gontijo S, Richardson M, et al. Coagulant thrombin-like enzyme (barnettobin) from Bothrops barnetti venom: Molecular sequence analysis of its cDNA and biochemical properties . Biochimie. 2013 Jul;95(7):1476-86. doi: 10.1016/j.biochi.2013.03.015.
8. Sandoval G, Lazo F, Rodriguez E, Yarlequé A, Zingali R. Identificación molecular y actividad sobre sustratos cromogénicos de la venombina A del veneno de la serpiente peruana Bothrops atrox. Rev Peru Biol. 2010;17(3): 365-70.
9. Valeriano-Zapana J, Segovia-Cruz FS, Rojas-Hualpa JM, Martins-de-Souza D, Ponce-Soto LA, Marangoni S. Functional and structural characterization of a new serine protease with thrombin-like activity TLBan from Bothrops andianus (Andean Lancehead) snake venom . Toxicon. 2012 Feb;59(2):231-40. doi: 10.1016/j.toxicon.2011.11.018.
10. Pirkle H. Thrombin-like enzymes from snake venoms: an updated inventory. Scientific and Standardization Committee’s Registry of Exogenous Hemostatic Factors. Thromb Haemost. 1998 Mar;79(3):675-83.
11. Olascoaga ME, Zavaleta A, Marsh NA. Preliminary studies of the effects of a Peruvian snake Bothrops pictus (jergon of the coast) venom upon fibrinogen. Toxicon. 1988;26(5):501-4.
12. Orejuela P, Zavaleta A, Salas M, Marsh N. Thrombin-like activity in snake venoms from Peruvian Bothrops and Lachesis genera. Toxicon. 1991;29(9):1151-4.
13. Rojas E, Quesada L, Arce V, Lomonte B, Rojas G Gutiérrez JM. Neutralization of four Peruvian Bothrops sp. snake venoms by polyvalent antivenoms produced in Perú and Costa Rica: preclinical assessment. Acta Trop. 2005 Jan;93(1):85-95.
14. Laing GD, Yarlequé A, Marcelo A, Rodriguez E, Warrell DA, Theakston RD. Preclinical testing of three South American antivenoms against the venoms of five medically-important Peruvian snake venoms. Toxicon. 2004 Jul; 44(1):103-6.
15. Maguiña C, Henriquéz C, Ilquimiche L, Mostorino R, Gotuzzo E, Legua P, et al. Ofidismo por Bothrops pictus en el Hospital Nacional Cayetano Heredia: Estudio prospectivo de 23 casos. Folia Dermatol Peru. 1998;9(1-2):41-8.
16. Mesía M, Lazo F, Yarlequé A. Purificación y caracterización de un nuevo principio coagulante del veneno de la serpiente peruana Bothrops pictus. Rev Soc Quím Perú. 2011;77(3):182-90.
17. Koh DC, Armugam A, Jeyseelan K. Snake venom components and their applications in biomedicine. Cell Mol Life Sci. 2006 Dec;63(24):3030-41.
18. Markland F, Swenson S. Venombin A. En: Barret A, Rawlings N, Woessner F, ed. Handbook of Proteolytic Enzymes. 2da ed. Vol 2. London: Elseiver Academis Press; 2004. p. 1715-22.
19. Magalhães A, Magalhães HP, Richardson M, Gontijo S, Ferreira RN, Almeida AP, et al. Purification and properties of a coagulant thrombin-like enzyme from the venom of Bothrops leucurus. Comp Biochem Physiol A Mol Integr Physiol. 2007 Apr;146(4):565-75.
20. Winzler R. Determinations of serum glycoproteins. En: Methods of Biochemical Analysis. New York: Interscience; 1955. p. 279-311.
21. Warren L.. The thiobarbituric acid assay of sialic acids. J Biol Chem. 1959 Aug;234(8):1971-5.
22. Instituto Cloromido Picado, Universidad de Costa Rica. Determinación de la actividad coagulante. Determinación de actividades tóxicas de venenos de serpientes y su neutralización por antivenenos. Manual de métodos de laboratorio. San José: Facultad de Microbiología; 2007. p. 21-2.
23. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970 Aug 15;227(5259):680-5.
24. Costa FL, Rodrigues RS, Izidoro LF, Menaldo DL, Hamaguchi A, Homsi-Brandeburgo MI, et al. Biochemical and functional properties of a thrombin-like enzyme isolated from Bothrops pauloensis snake venom. Toxicon. 2009 Nov;54(6):725-35. doi: 10.1016/j.toxicon.2009.05.040.
Correspondencia: Dan Erick Vivas Ruiz,
Dirección: Laboratorio de Biología Molecular (Laboratorio 120), Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Av. Venezuela/Av. Universitaria Cercado de Lima, Perú.
Teléfono: 016197000 anexo 1558.
Correo electrónico: dvivasr@unmsm.edu.pe
Recibido: 15-01-15
Aprobado: 17-06-15