Maribel Riveros
Maria Pinedo-Bardales
Joaquim Ruiz
Theresa J. Ochoa
10.17843/rpmesp.2021.381.5750
ORIGINAL BREVE
Frecuencia de genes que codifican proteínas autotransportadoras serin-proteasa de Enterobacteriaceae (SPATE) en cepas de Escherichia coli difusamente adherente (DAEC) provenientes de niños con y sin diarrea
Frequency of serine protease autotransporters of Enterobacteriaceae (SPATE) encoding genes in diffusely adherent Escherichia coli (DAEC) isolates from children with and without diarrea
Karen Ocampo ![]() 1,2, Bióloga
1,2, Bióloga
Maribel
Riveros ![]() 1,3,
magíster en Biología Molecular
1,3,
magíster en Biología Molecular
Maria
Pinedo-Bardales ![]() 1,4, estudiante de Biología
1,4, estudiante de Biología
Joaquim Ruiz ![]() 5,6, microbiólogo, doctor en Biología
5,6, microbiólogo, doctor en Biología
Theresa J.
Ochoa ![]() 1,7,médica especialista en Infectología Pediátrica
1,7,médica especialista en Infectología Pediátrica
1 Laboratorio de Infectología Pediátrica, Instituto de Medicina
Tropical «Alexander von Humboldt», Universidad Peruana Cayetano Heredia,
Lima, Perú.
2 Universidad Nacional Federico Villarreal, Lima, Perú.
3 Facultad de Medicina, Universidad Peruana Cayetano Heredia, Lima,
Perú
4 Facultad de Ciencias, Universidad Nacional de Piura, Piura, Perú.
5 ISGlobal, Hospital Clínic - Universitat de Barcelona, Barcelona,
España.
6 Laboratorio de Microbiología Molecular y Genómica Bacteriana,
Universidad Científica del Sur, Lima, Perú.
7 School of Public Health, University of Texas Health Science Center
at Houston, Houston, Estados Unidos.
El presente estudio forma parte de la tesis de Ocampo K., Frecuencia de las proteínas Autotransportadoras Serin-Proteasa de Enterobacteriaceae (SPATE) en cepas de Escherichia coli difusamente adherente (DAEC) aisladas de niños con y sin diarrea [Tesis de Licenciatura]. Lima: Facultad de Ciencias Naturales y Matemática, Universidad Nacional Federico Villarreal; 2015.
RESUMEN
El objetivo del estudio fue determinar la frecuencia de seis genes que codifican proteínas autotransportadoras serin-proteasa de Enterobacteriaceae (SPATE) en aislamientos de Escherichia coli difusamente adherente (DAEC) provenientes de niños con diarrea (NCD, n=63) y sin diarrea (NSD, n=41) de Lima, Perú. Los NSD se consideraron como grupo control. Para la detección de los genes se estandarizaron 2 PCRs múltiples: triple A (sigA, pet, espP) y triple B (sat, pic, espC). En ambos grupos el gen SPATE más frecuente fue sat (39,7% de NCD y 41,5% de NSD), seguido de espP (20,6% y 9,7% en NCD y NSD respectivamente). Los otros genes se detectaron en proporciones inferiores al 10,0%, en el siguiente orden de frecuencia: pet, sigA, espC y pic, sin diferencias significativas entre los grupos. Se concluye que Sat es la SPATE más frecuente en cepas DAEC, y que estas cepas pueden poseer genes SPATE independientemente de si se aíslan en NCD o NSD.
Palabras clave: Escherichia coli de Adherencia Difusa; Factores de virulencia; Diarrea; Niño (fuente: DeCS BIREME).
ABSTRACT
The aim of the study was to determine the frequency of six genes encoding serine protease autotransporter proteins Enterobacteriaceae (SPATE) in diffusely adherent Escherichia coli (DAEC) isolates from children with (WD, n=63) and without diarrhea (WOD, n=41) from Lima, Peru. WOD were considered a control group. For the detection of the genes, 2 multiple PCRs were standardized: triple A (sigA, pet, espP) and triple B (sat, pic, espC). In both groups, the most frequent SPATE gene was Sat (39.7% of WD and 41.5% of WOD), followed by spP (20.6% and 9.7% in WD and WOD respectively). The other genes were detected in proportions lower than 10.0%, in the following order of frequency: pet, sigA, espC and pic, without significant differences between the groups. It was concluded that Sat is the most frequent SPATE in DAEC and that these strains may possess SPATE genes regardless of whether they are isolated in WD or WOD.
Keywords: Diffusely Adherent Escherichia coli; Virulence Factors; Diarrhea; Child (source: MeSH NLM).
INTRODUCCIÓN
La diarrea es una de las principales causas de muerte en niños menores de cinco años en el mundo. En este grupo etario, Escherichia coli es uno de los agentes etiológicos más frecuentes (1). Según su patogénesis, las E. coli diarreogénicas (DEC) se dividen en seis patotipos: enteroagregativa (EAEC), enterotoxigénica (ETEC), enteropatógena (EPEC), verotoxigénica (VTEC), enteroinvasiva (EIEC) y difusamente adherente (DAEC) (2).
Las DAEC constituyen un grupo heterogéneo y se han asociado particularmente con diarrea persistente en los países en desarrollo, afectando principalmente a niños mayores de un año (2). En Perú, se ha descrito DAEC en muestras de niños menores de cinco años con diarrea persistente con frecuencias de 15,0% y 17,0% para Lima y Lambayeque respectivamente (3,4). Además, se ha reportado que DAEC presenta diferencias en los patrones de adherencia, polimerización de actina y motilidad dependiendo de sí se aísla de niños con diarrea o asintomáticos, y se sugiere la determinación de los factores de virulencia presentes en estas cepas (5).
Las cepas patógenas de E. coli, tanto extraintestinales como DEC, se caracterizan por poseer factores de virulencia, los cuales les ayudan a superar las defensas del huésped y colonizar o invadir el tracto urinario (cepas uropatógenas) o gastrointestinal (cepas DEC) (6,7). Los factores de virulencia incluyen toxinas, adhesinas y otras enzimas hidrolíticas secretadas a través de vías conocidas como sistemas de secreción de proteínas (8). Estas vías se clasifican en cinco grupos principales: tipo I, II, III, IV y autotransportadoras. Dentro del grupo de proteínas autotransportadoras, la subclase más frecuente está conformada por las autotransportadoras de serin-proteasa de Enterobacteriaceae (SPATE, por sus siglas en inglés), las cuales se han identificado en bacterias gramnegativas como Salmonella y Shigella (9,10).
En las DEC, la SPATE más reportada es la toxina autotransportadora secretada (sat) (2,11,12). Aunque la presencia de SPATEs en cepas DAEC se ha evaluado escasamente, se ha reportado que sat es la más frecuente, encontrándose en un mayor porcentaje en cepas aisladas de pacientes con diarrea que en aquellas de portadores asintomáticos (12). La escasa evidencia científica sobre las principales SPATEs presentes en cepas DAEC hace que sea relevante analizar su frecuencia en este patotipo. Por lo tanto, el objetivo del estudio fue determinar la frecuencia de seis genes que codifican SPATE en aislamientos de DAEC de niños con y sin diarrea.
|
MENSAJES CLAVE |
|
Motivación para realizar el estudio: Es necesario evaluar la presencia de proteínas autotransportadoras serin-proteasa de Enterobacteriaceae (SPATE) en cepas de Escherichia coli difusamente adherente (DAEC), debido a que estas proteínas facilitan la invasión del huésped durante la infección bacteriana. Principales hallazgos: Se pueden encontrar genes que codifican SPATE en cepas DAEC, tanto en muestras de pacientes con diarrea como en niños sanos. El gen sat es la SPATE más frecuente en estas cepas. Implicancias: Conocer los genes que codifican los mecanismos moleculares involucrados en la interacción del ser humano y el agente infeccioso, permitirá orientar investigaciones dirigidas a pruebas de diagnóstico y control o el desarrollo de futuros candidatos a vacunas. |
EL ESTUDIO
Cepas
Se analizaron 104 cepas aisladas de niños menores de cinco años con diarrea (n=63) y asintomáticos, los cuales se consideraron como grupo control (n=41). Los aislamientos provenían de tres estudios de cohortes realizados en el distrito de Lima (Perú) entre 2006 y 2010, siendo identificados molecularmente como DAEC mediante la detección del gen daaD (13) y reportados por Ochoa et al. (3). Las cepas DAEC se reactivaron en agar MacConkey y se incubaron durante 24 horas a 37 ºC. Posteriormente, se extrajo el ADN mediante el método de choque térmico.
Los proyectos de los cuales se obtuvieron las cepas incluidas en el estudio (Códigos SIDISI: 50744, 57025 y 50908) fueron aprobados por el Comité de Ética de la Universidad Peruana Cayetano Heredia.
Detección de SPATE
La presencia de SPATE se determinó por medio de dos reacciones en cadena de la polimerasa (PCR) múltiples usando los cebadores reportados en la Tabla 1. Inicialmente, se estandarizaron PCR simples para la detección individual de cada gen (Tabla 1) para establecer las condiciones óptimas.
Tabla 1. Lista de cebadores utilizados en el estudio.
|
Técnica |
Gen |
Primer |
Pares de bases |
Referencia |
|
PCR triple A |
espP |
Forward: GTCCATGCAGGGACATGCCA |
547 |
9 |
|
Reverse: TCACATCAGCACCGTTCTCTAT |
||||
|
pet |
Forward: GGCACAGAAT AAAGGGGTGTTT |
302 |
9 |
|
|
Reverse: CCTCTTGTTTCCACGACATAC |
||||
|
sigA |
Forward: CCGACTTCTCACTTTCTCCCG |
430 |
9 |
|
|
Reverse: CCATCCAGCTGCATAGTGTTTG |
||||
|
PCR triple B |
sat |
Forward: TCAGAAGCTCAGCGAATCATTG |
930 |
9 |
|
Reverse: CCATTATCACCAGTAAAACGCACC |
||||
|
espC |
Forward: TAGTGCAGTGCAGAAAGCAGTT |
301 |
8 |
|
|
Reverse: AGTTTTCCTGTTGCTGTATGCC |
||||
|
pic |
Forward: ACTGGATCTTAAGGCTCAGGAT |
572 |
9 |
|
|
Reverse: GACTTAATGTCACTGTTCAGCG |
Reverse: GACTTAATGTCACTGTTCAGCG
PCR: reacción en cadena de la polimerasa.
Tras evaluar la efectividad de la amplificación de los productos a temperaturas adecuadas, se procedió al diseño de PCR múltiples y la selección de la mejor combinación de cebadores. Para ello, se utilizó la página web de Integrated DNA Technologies (http://www.idtdna.com), donde se evaluó el contenido de guanina y citocina (% GC), la temperatura de Melting (Tm), la formación de horquillas y los dímeros generados. Posteriormente, se analizó la formación de estructuras secundarias con el ADN diana en mfold (http://mfold.rna.albany.edu/?q=mfold/dna-folding-from).
Se obtuvieron bandas específicas, definidas y diferenciables entre sí (Figura 1A y 1B), al realizar la amplificación múltiple de pet, sigA y espP (triple A), y la de sat, espC y pic (triple B).
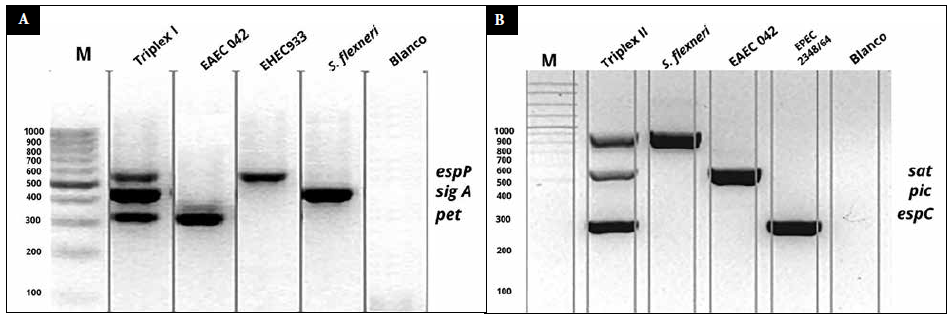
Figura 1. A. Gel de agarosa al 2%, el segundo carril corresponde al triplex A, los carriles 3, 4 y 5 corresponde a los genes individuales, espP (547 pb), sigA (430 pb), pet (302 pb), temperatura de hibridación de 60 °C, blanco (control negativo). B. Gel de agarosa al 2%, el segundo carril corresponde al triplex B, los carriles 3, 4 y 5 corresponde a los genes individuales, sat (930 pb), pic (572 pb), espC (320 pb), temperatura de hibridación de 64 °C, blanco (control negativo).
Para estandarizar el triple A se evaluó el gradiente de temperatura de hibridación de 54 a 64 °C a un volumen final de 25 µl que contenía: dNTP (0,23 mM), MgCl2 (1,7 µM), cebadores pet y sigA a 0,5 µM, cebadores espP a 0,6 µM, Taq buffer (concentración final 1X) y GoTaq DNA Polimerasa (Promega, Francia) (5U/µM); usando las siguientes condiciones: desnaturalización inicial a 94 °C por cinco minutos y 30 ciclos con desnaturalización 94 °C por un minuto, hibridación por 30 segundos, elongación 72 °C por un minuto, y una elongación final a 72 °C por cinco minutos. Bajo estas condiciones, el triple A presentó una mejor definición de bandas en la temperatura de hibridación de 60 °C.
En la estandarización del triple B, se evaluaron las temperaturas de hibridación de 54 a 64 °C, a un volumen final de 25 µl que contenía: dNTP (0,08 mM), MgCl2 (1,7 µM), cebadores (espC, pic y sat) a 0,5 µM, Taq buffer (concentración final 1X) y Taq (5U/µM); bajo las siguientes condiciones: desnaturalización inicial a 94 °C por cinco minutos y 30 ciclos con desnaturalización 95°C por un minuto, hibridación por un minuto, elongación 72 °C por un minuto, y una elongación final a 72 °C por diez minutos. Se observó una mayor definición de los productos de PCR en la temperatura de 64 °C.
Las reacciones fueron realizadas en el Termociclador Applied Biosystems Veriti 96-Well Thermal Cycler (Fisher Scientific, Estados Unidos). El control negativo fue agua estéril y los controles positivos fueron las cepas de E. coli EAEC 042 para pet y pic, EHEC 933 para espP y EPEC 2348/64 para espC, y la cepa de Shigella flexneri 1106 para sigA y sat. Las cepas control se obtuvieron de la colección interna del Laboratorio de Infectología Pediátrica del Instituto de Medicina Tropical «Alexander von Humboldt».
Los productos de PCR fueron visualizados en geles de agarosa al 2%, teñidos con bromuro de etidio a 90V por 80 minutos.
Análisis de datos
La frecuencia y comparación de todas las variables se realizó mediante la prueba de Chi cuadrado utilizando el programa Epi- Info 7. El programa GenAlEx fue empleado en el análisis de perfiles genómicos.
HALLAZGOS
De los 104 aislamientos incluidos en el estudio, 58,6% presentaron al menos un gen codificante para SPATEs.
El gen más frecuente fue sat (40,4%), en proporciones similares en aislados de niños con diarrea (39,7%) y asintomáticos (41,5%); seguido por espP (16,3%), sigA (10,0%) y pet (7,0%). Los genes pic y espC presentaron una frecuencia de 1,9%. El gen espP se encontró asociado a muestras de casos diarreicos en un 20,6%, mientras que en controles (aislamientos de niños asintomáticos) se detectó en 9,7%, aunque sin diferencias significativas (Tabla 2).
Tabla 2. Frecuencias de genes SPATE en cepas DAEC de niños con y sin diarrea menores de un año.
|
Característica |
Genes SPATE |
|||||
|
pet |
sigA |
espP |
sat |
pic |
espC |
|
|
NCD (n=63) |
5 (7,9%) |
5 (7,9%) |
13 (20,6%) |
25 (39,7%) |
0 |
0 |
|
NSD (n=41) |
2 (4,9%) |
5 (12,2%) |
4 (9,7%) |
17 (41,5%) |
2 (4,9%) |
2 (4,9%) |
NCD: niños con diarrea; NSD: niños sin diarrea.
Por otro lado, no se observó asociación entre la presencia del resto de SPATEs y el origen (ya sea caso o control) de las cepas DAEC.
Se identificaron 12 perfiles genéticos, determinados por las combinaciones de genes codificantes para SPATEs (Tabla 3). El perfil más común entre las muestras diarreicas fue la ausencia de SPATEs (41,4%), seguido por la presencia solo del gen sat (30,8%). Los nueve perfiles restantes fueron menos frecuentes sin que ninguno sobrepasase los diez aislamientos. Ninguno de los perfiles se asoció con la presencia o ausencia de diarrea.
Tabla 3. Perfil genético de cepas DAEC aisladas de niños con y sin diarrea.
|
Perfiles |
pet |
espP |
sigA |
sat |
pic |
espC |
NCD (n=63) |
NSD (n=41) |
|
1 |
|
|
|
|
|
|
26 |
17 |
|
2 |
|
|
|
+ |
|
|
20 |
12 |
|
3 |
|
+ |
|
|
|
|
6 |
2 |
|
4 |
|
+ |
|
+ |
|
|
4 |
1 |
|
5 |
+ |
|
+ |
|
|
|
2 |
0 |
|
6 |
|
|
+ |
|
|
|
2 |
2 |
|
7 |
+ |
|
+ |
|
|
+ |
1 |
0 |
|
8 |
+ |
+ |
|
+ |
|
|
1 |
0 |
|
9 |
+ |
+ |
|
|
|
|
1 |
1 |
|
10 |
+ |
|
|
+ |
|
|
0 |
1 |
|
11 |
|
|
+ |
+ |
|
|
0 |
3 |
|
12 |
|
|
|
|
|
+ |
0 |
2 |
NCD: niños con diarrea; NSD: niños sin diarrea; +: presencia del gen.
DISCUSIÓN
En las muestras analizadas, el gen sat presentó la frecuencia más alta, tanto en casos (39,7%) como en controles (41,5%), resultado similar al reporte de Taddei et al. (11), que encontraron un 44,0% de positividad para este gen tras analizar 146 cepas DAEC. De igual forma, Boisen et al. (9) detectaron el gen sat en el 30,0% de cepas DAEC analizadas. Aunque, a diferencia de lo reportado por otros autores (12), la frecuencia de sat fue similar en casos y controles. Mansan et al. observaron que la expresión de este gen es capaz de causar lesiones en uniones estrechas de células epiteliales, conduciendo a un aumento de la permeabilidad y pérdida de líquido, así como, que la presencia del gen sat se asociaba a casos de diarrea (46,0% vs. 19,0%; valor de p<0,05) (14). En el presente estudio, no se encontró esta asociación; sin embargo, los resultados concuerdan con la investigación de Toloza et al. (15), donde concluyeron que el papel de sat como factor de virulencia depende de la carga genética de las cepas. Con respecto a esto, se debe tener en cuenta la posibilidad de alteraciones a nivel de expresión del gen, mutaciones puntuales que afecten a su funcionalidad y la carga bacteriana, que en el caso de otras cepas DEC (las EPEC) se ha relacionado directamente con la presencia/ausencia de síntomas (16).
El segundo gen más frecuente fue espP, el cual se encuentra fuertemente asociado con EHEC y EPEC atípicas, siendo reportado con menor frecuencia en otras DEC, como es el caso de EAEC o EPEC típicas. El gen espP codifica una proteína capaz de degradar el factor V de coagulación, participando así en la producción del síndrome urémico-hemolítico, característico de EHEC. Asimismo, es responsable de cortar el factor de complemento C3/C3b, lo cual facilita la colonización del intestino (10).
El gen sigA se presentó con una frecuencia del 10,0%, similar a lo descrito por Boisen et al. (9). Este gen fue reportado inicialmente en la isla de patogenicidad SHI-1 presente en S. flexneri 2a (17), sin embargo, la detección de este gen en ausencia de otros componentes de esta isla de patogenicidad sugiere su presencia en otras estructuras genéticas (9).
Los tres genes restantes se detectaron en frecuencias inferiores al 10%. El gen pet se detectó en 7,0% de las cepas, en su mayoría provenientes de niños con diarrea. En otras DEC, como EPEC y EAEC, pet ha sido reportado con frecuencias entre el 5 y el 20,0% respectivamente (7,9,18). El pet es capaz de producir efectos enterotoxigénicos y citopáticos, y posee la capacidad de degradar fodrina, lo cual produce la desorganización de la actina del citoesqueleto celular (19).
Tanto pic como espC se encontraron con frecuencias menores al 2,0% y solo en cepas DAEC provenientes de niños asintomáticos. espC ha sido previamente descrito en cepas EPEC (17). La proteína que codifica este gen desempeña un papel relevante en la muerte celular inducida por EPEC, regulando la secreción de factores de virulencia como EspA o EspD. pic se ha reportado en otras DEC, como EAEC y UPEC (20).
El análisis de la presencia concomitante de diferentes SPATE mostró la presencia de 12 perfiles de asociación de genes diferentes. No obstante, ninguno de los patrones observados se pudo relacionar con casos o controles. Esta no asociación puede deberse a varios factores, incluyendo la presencia de diferentes variantes alélicas o los niveles de expresión de los genes evaluados (16).
Una limitación de este estudio es que, debido a la ausencia de datos clínico de los pacientes, no fue posible realizar un análisis más complejo ni relacionarlo con otros síntomas. Asimismo, el número de aislamientos disponibles puede influir en la no detección de diferencia significativa entre los grupos caso y control. Sin embargo, este estudio presenta la primera descripción de genes SPATEs en cepas DAEC aisladas de niños peruanos.
En conclusión, 58,6% de las cepas DAEC aisladas de niños con y sin diarrea presentaron al menos un gen codificante de SPATE. En ambos grupos, sat y espP fueron los genes más frecuentes. En general, los resultados mostraron una heterogeneidad a nivel de SPATEs en cepas DAEC, por ello se sugiere realizar estudios adicionales haciendo uso de un mayor número de aislamientos, tanto de niños con diarrea como asintomáticos. La detección de SPATEs es importante debido a que estas proteínas facilitan la invasión del huésped durante la infección bacteriana y algunos genes codificantes (como pet y espP) pueden ser transmitidos mediante plásmidos a otras especies bacterianas.
Agradecimientos:
ISGlobal es miembro del programa CERCA, Generalitat de Catalunya.
REFERENCIAS BIBLIOGRÁFICAS
1. Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet. 2013;382:209–22. doi: 10.1016/S0140-6736(13)60844-2.
2. Servin AL. Pathogenesis of human diffusely adhering Escherichia coli expressing Afa/Dr adhesins (Afa/Dr DAEC): Current insights and future challenges. Clin Microbiol Rev. 2014;27:823–69. doi: 10.1128/CMR.00036-14.
3. Ochoa TJ, Mercado EH, Durand D, Rivera FP, Mosquito S, Contreras C, et al. Frecuencia y patotipos de Escherichia coli diarrogénica en niños peruanos con y sin diarrea. Rev Peru Med Exp Salud Publica. 2011;28(1):13-20. doi: 10.1590/s1726-46342011000100003.
4. Yacarini-Martínez AE, Arriaga-Deza EV, Alvarado-Pineda RL, Fupuy-Chung JA. Genes de virulencia de Escherichia coli detectados en muestras diarreicas de niños de la Región Lambayeque - Perú. Horiz Med (Lima). 2019;19(1):7-12. doi: 10.24265/horizmed.2019.v19n1.02.
5. Riveros M, Barletta F, Cabello M, Durand D, Mercado EH, Contreras C, et al. Patrones de adherencia de cepas de Escherichia coli Difusamente adherente (DAEC) provenientes de niños con y sin diarrea. Rev Peru Med Exp Salud Publica. 2011;28(1):21-8. doi: 10.1590/s1726-46342011000100004.
6. Johnson JR. Virulence factors in Escherichia coli urinary tract infection. Clin Microbiol Rev. 1991;4:80–128. doi: 10.1128/cmr.4.1.80.
7. Durand D, Contreras CA, Mosquito S, Ruíz J, Cleary TG, Ochoa TJ. pic gene of enteroaggregative Escherichia coli and its association with diarrhea in Peruvian children. Pathog Dis. 2016;74:ftw054. doi: 10.1093/femspd/ftw054.
8. Restieri C, Garriss G, Locas M-C, Dozois CM. Autotransporter-encoding sequences are phylogenetically distributed among Escherichia coli clinical isolates and reference strains. Appl Environ Microbiol. 2007;73(5):1553–62. doi: 10.1128/AEM.01542-06.
9. Boisen N, Ruiz-Perez F, Scheutz F, Krogfelt KA, Nataro JP. High prevalence of serine protease autotransporter cytotoxins among strains of enteroaggregative Escherichia coli. Am J Trop Med Hyg. 2009;80(2):294–301. doi: 10.4269/ajtmh.2009.80.294.
10. Weiss A, Brockmeyer J. Prevalence, biogenesis, and functionality of the serine protease autotransporter. EspP Toxins (Basel). 2012;5(1):25-48. doi: 10.3390/toxins5010025.
11. Taddei CR, Moreno ACR, Fernandes Filho A, Montemor LP., Martinez MB. Prevalence of secreted autotransporter toxin gene among diffusely adhering Escherichia coli isolated from stools of children. FEMS Microbiol Lett. 2003;227:249–53. doi: 10.1016/S0378-1097(03)00688-8.
12. Guignot J, Chaplais C, Coconnier-Polter MH, Servin AL. The secreted autotransporter toxin, Sat, functions as a virulence factor in Afa/Dr diffusely adhering Escherichia coli by promoting lesions in tight junction of polarized epithelial cells. Cell Microbiol. 2007;9(1):204-21. doi: 10.1111/j.1462-5822.2006.00782.x.
13. Guion CE, Ochoa TJ, Walker CM, Barletta F, Cleary TG. Detection of Diarrheagenic Escherichia coli by use of melting-curve analysis and real-time multiplex PCR. J Clin Microbiol. 2008;46:1752–7. doi: 10.1128/JCM.02341-07.
14. Mansan-Almeida R, Pereira A, Giugliano L. Diffusely adherent Escherichia coli strains isolated from children and adults constitute two different populations. BMC Microbiol. 2013;13:22. doi: 10.1186/1471-2180-13-22.
15. Toloza L, Giménez R, Fábrega MJ, Alvarez CS, Aguilera L, Cañas MA, et al. The secreted autotransporter toxin (Sat) does not act as a virulence factor in the probiotic Escherichia coli strain Nissle 1917. BMC Microbiol. 2015;15:250. doi: 10.1186/s12866-015-0591-5.
16. Barletta F, Ochoa TJ, Cleary TG. Multiplex Real-Time PCR (MRT-PCR) for diarrheagenic. Methods Mol Biol. 2013;943: 307-14. doi: 10.1093/cid/cir730.
17. Al-Hasani K, Henderson IR, Sakellaris H, Rajakumar K, Grant T, Nataro JP, et al. The sigA gene which is borne on the she pathogenicity island of Shigella flexneri 2a encodes an exported cytopathic protease involved in intestinal fluid accumulation. Infect Immun. 2000;68(5):2457–63. doi: 10.1128/iai.68.5.2457-2463.2000.
18. Abreu AG, Bueris V, Porangaba TM, Sircili MP, Navarro-Garcia F, Elias WP. Autotransporter protein-encoding genes of diarrheagenic Escherichia coli are found in both typical and atypical enteropathogenic E. coli strains. Appl Environ Microbiol. 2013;79(1):411–4. doi: 10.1128/AEM.02635-12.
19. Navarro-Garcia F, Sonnested M, Teter K. Host-Toxin interactions involving EspC and Pet, two serine protease autotransporters of the Enterobacteriaceae. Toxins (Basel). 2010;2(5):1134–47. doi: 10.3390/toxins2051134.
20. Pokharel P, Habouria H, Bessaiah H, Dozois CM. Serine protease autotransporters of the Enterobacteriaceae (SPATEs): out and about and chopping it up. Microorganisms. 2019;7(12):594. doi: 10.3390/microorganisms7120594.
Financiamiento: Esta investigación fue apoyada por la Agencia Española de Cooperación Internacional y Desarrollo (AECID) (D/030509/10). Joaquim Ruiz fue financiado por el programa I3 del Instituto de Salud Carlos III (número de concesión: CES11/012) y por el Fondo Nacional de Desarrollo Científico, Tecnológico y de Innovación Tecnológica (FONDECYT - Perú) en el marco del «Proyecto de Mejoramiento y Ampliación de los Servicios del Sistema Nacional de Ciencia, Tecnología e Innovación Tecnológica» (número de contrato 08-2019-FONDECYT-BM-INC-INV).
Material suplementario: Disponible en la versión electrónica de la RPMESP.
Citar como: Ocampo K, Riveros M, Pinedo-Bardales M, Ruiz J, Ochoa TJ. Frecuencia de genes que codifican proteínas autotransportadoras serin-proteasa de Enterobacteriaceae (SPATE) en cepas de Escherichia coli difusamente adherente (DAEC) provenientes de niños con y sin diarrea. Rev Peru Med Exp Salud Publica. 2021;38(1):124-9. doi: https://doi.org/10.17843/rpmesp.2021.381.5750.
Correspondencia: Maribel Riveros; Universidad Peruana Cayetano Heredia, Honorio Delgado 430, San Martín de Porres, Lima, Perú; maribel.riveros@upch.pe
Contribuciones de autoría: KO y MR participaron en la recolección de resultados, análisis e interpretación de datos y redacción del artículo. MP realizó el análisis e interpretación de datos y redacción del artículo. JR y TJO participaron en la concepción y diseño del artículo, obtención de financiamiento, redacción del artículo y revisión crítica. Todos los autores aprobaron la versión final y asumen responsabilidad de su contenido.
Conflictos de interés: Los autores no declaran conflictos de interés.
Recibido: 06/05/2020
Aprobado: 30/09/2020
En línea: 24/02/2021